

第八节. 化石及地球的年龄
进化论的假说要成立,很关键的一点是需要经过漫长的时间,好让自然选择对缓慢积累的遗传变异起作用。这篇文章到这里为止,为了使前面的讨论简化,都有意回避了一个问题,那就是化石形成年代的测定。在前述中,我们经常会看到如某一化石经测定为1.5亿年前形成的。然而我们在这一节首先要指出的是,其实很多化石的年代测定是有问题的,那些化石形成的年代到现在其实很有可能并没有那么久远。换句话说,某一被认为是1.5亿年前的化石很有可能并不是在那么久之前形成的,这样进化论所要求的“漫长的时间”很有可能是没法满足的。
现代科技最常用的年代测算方法就是放射性年代测定法(Radiometric Dating)。它是利用测定被测物中某些放射性母元素与其衰变(Radioactive Decay)产物子元素的比率,之后应用这种放射性母元素的半衰期(Half-life)计算年代的方法。按照这种方法,理论上来说,样本中子元素对母元素的比率越高,年代就应该越久远。这就好比一个沙漏,其上部的沙粒(母元素物质)通过恒定的速率向下部运动而致使出现下部的沙粒(子元素物质),通过对上下部分沙粒的数量的观察,就可得到时间的概念。
最常用的放射性测定法包括:
-> 碳-14测年法 (多用于测量年代较近的生物遗迹)
-> 钾氩测年法 (多用于测量被认为年代较近的岩浆岩)
-> 铀系测年法 (多用于测量被认为年代较远的岩浆岩)
其中碳-14测年法的原理是:生物体在活着的时候会因呼吸、进食等不断地从外界摄入碳-14(一种地球环境中的放射性元素,可衰变为氮-14,其半衰期约为5700年),最终体内碳-14与碳-12的比值会达到与环境一致(现今观察该比值基本不变);当生物体死亡时,碳-14的摄入停止,之后因其遗体中碳-14的衰变而使其中的碳-14与碳-12的比值发生变化,在假设环境中碳-14与碳-12比值在地球历史中从未大幅改变的前提下,理论上通过测定碳-14与碳-12的比值就可以测算该生物的死亡年代。不过碳-14的半衰期比较短,它会以较快的速度衰变为氮-14,因此碳-14测年法的应用只局限于5到6万年(现代测量仪器只能检测到的衰变时间不超过10万年内遗留下的碳-14)。碳-14测年法经过树木年轮校正(这里也有不确定因素,因为很多树木并非严格地一年生长出一圈年轮),据称对于被认为1万年以内的测年结果误差可控制在60年以内,被认为3万年以内的结果误差为350年以内。在离我们较近的年代内,特别是几千年内(注意迄今为止地球上发现的最古老的树木年轮只局限在约5000轮的范围内),碳-14测年法应该是具有一定的可信度的。
而钾氩测年法现在用于测量被认为超过10万年前形成的岩石,铀系测年法多用于一般是被认为0.5至50万年的岩石。
必须清楚的是,使用放射性测年法是建立在如下假设的基础上:
-> 在岩石形成时里面没有子元素(或母子元素初始含量比率已知,如碳-14测年法的情况);
-> 母元素衰变速率即半衰期(按现今测量值)恒定;
-> 岩石自形成后一直是封闭系统,即没有加入新的母元素和子元素,这些元素也不曾从岩石中流失。
在学界,放射性测年法在二十世纪初期被发明以来,发展出一系列的复杂的测试方法和仪器,它看似言之凿凿,却经常给出不一致甚至完全矛盾的年代结论,这是一个很多学者心知肚明的秘密。比如有学者就抱着“如果测年结果支持我们的理论,我们就会把它放在正文中;如果它并不完全与我们的理论相矛盾,我们就会把它放在脚注中;如果它完全与期望相悖,我们就会把它删除”的态度。
到底是什么造成了放射性测年的众多方法经常给出不一致甚至完全矛盾的年代结论呢?为了弄明白这个问题,美国的”创造研究所”(Institute for Creation Research) 组织七位科学家,使用独立的实验室,进行了一个为期八年(1997~2005)的名为”放射性同位素与地球年龄”(Radioisotopes and the Age of the Earth /RATE)的项目。这些科学家中包括地质学家、物理学家、地球物理学家、气象学家等。RATE项目的研究成果如论文、总结、影音等现在被保存在网址 https://www.icr.org/rate/。
RATE项目发现,放射性测年法所必须的上述三条假设在实际应用中都可能出现问题。比如物理学家已经注意到某些放射性母元素其半衰期会随时间变化,如某些铀同位素衰变为铅同位素的速率可能不是恒定的,在过去比在现在快很多,至今确切机制不明;而起始时刻不存在子元素(极少量的初始子元素对年轻岩石测定会产生极大的误差,会让岩石看起来古老的多),及系统一直维持封闭,这些在自然界中不可能完全保证。由此,当有其它条件所限或测量的年代被认为久远,而不能使用在几千年范围内较准确的碳-14测年法时,由其它测年法测得的结果有可能比实际情况在年代上古老很多。许多观察都能印证这一点。比如岩石和其中的化石常常被测出互相十分矛盾的年龄;再比如实际上已知年龄的、就在我们眼前形成年轻火山熔岩被测定出有数百万年的岩龄(RATE指出有证据显示岩浆会从地幔中带来额外的子元素如氩-40);再比如被认为有15亿年的锆石晶体Zircon)中氦气(Helium)的存在(已知作为2号元素且为惰性气体的氦原子在锆石晶体的晶格中无法被极长时间封存而不逃逸);再比如在被认为有几亿年年龄的钻石里测出了碳-14(别忘了只有10万年内残留的碳-14才能被最精密的仪器测出)。
RATE项目推测由于放射性元素衰变速率在地球历史上曾发生大幅加速,因此为依赖半衰期为恒定值(也即各放射性元素现今测得值)的所谓能测量古老年代的钾氩测年法和铀系测年法等引入整体性误差,使得其实很年轻的样本被测算出被认为是很古老的年代。当样本的采纳比较严谨并且在实验条件严格的条件下,大量的钾氩和铀系这样的放射性测年法实验确实能给出趋势一致的年代范围,然而这并不能说明它们能够给出真正的较准确的绝对的年龄。这就好比在枪支射击准确性实验中,一把被机械固定的步枪总能打到靶子上范围很小的区域,但这个局部现象一定能说明这把步枪没有问题吗?如果我们意识到这把步枪的准星就不正、即它的前提就不正确,那它每次都打在最低环数的同一相近区域就不足为奇了。
进而我们要指出的是,其实不只是化石的年龄,地球的年龄都极有可能没有现在主流研究宣称的46亿年那么古老。测量地球的年龄其中很大部分使用的也是岩石的放射性测年法,比如主流学界测出地球上某些最古老的岩石其年龄为约46亿年,基于前面对测量久远年代的测年法可能给出不准确结论的介绍,这个46亿年只是主流学界基于现有的理论和方法而进行的、可能存在很大误差的推测,而且我们已经分析了它的可能的错误来源。其实就算测出的46亿年没有大错特错,这就能说明地球是46亿年前形成的吗?笔者认为很难——这个46亿年只是地球的组成物质的大概年龄,并不是地球的年龄,地球特别是地表的形成可以是在它的组成物质具备很久以后,这就像用面粉形成的时间点不能推测出面团形成的时间点一样。
如果这里的分析都基本正确的话,古老的化石就可能不是那么古老,地球也不像我们现在认为的那么、那么古老,面对一个年轻的“自然母亲”,则深陷困境的进化论所要克服的困难可能就又要多了一重。
第九节. 进化论的“进化”
达尔文主义的进化论从被提出到现在的超过一个半世纪的时间里,经历了几次大的变身和分身,现代所说的进化论(尤其是某些分身理论)可以说已经是与传统的达尔文主义在很多方面相去甚远了。很多进化论者可能觉得这也没什么,科学理论从来不都是不断发展、逐步完善的吗?看看牛顿经典的万有引力和力学定律,不也是被爱因斯坦的相对论指出只是在低速相对运动下的近似吗?这并不妨碍牛顿的伟大呀?是啊,那并不妨碍牛顿的伟大,因为他提出的万有引力和经典力学在人类生活的时空尺度上显示了强效的解释力和预测力,这一点从牛顿理论的诞生到今天从未改变过,更何况牛顿还有在数学和光学等领域的杰出贡献呢。相对论并没有否定经典牛顿理论在限定范围内的解释力和预测力。
那么安葬在牛顿墓旁的达尔文呢?我们已经看到他的进化理论从诞生到现在并没有真正极具信服力地解释生物物种的起源,他的理论也不曾准确地预测过什么,他在他书中的预测至今来看无一例外都破产了。我们也有极大的理由相信传统达尔文进化论以后所有的变身或分身理论,也都不能解释什么和预测什么,和传统进化论一样,它们充其量只能被称为假说(第十节详细讨论)。任何从达尔文肇始的基于遗传变异和自然选择的进化理论从未能真正地解释进化的机制,无论达尔文的衣钵继承者们如何绞尽脑汁,提出各种叠加在名词之上的重重的新名词,和各种基于假设之上的层层的新假设,他们理论的解释力总是显得那么的苍白与惨淡,更无需多言其理论的预测力了。
以下是对进化论原身,和有主要影响力的变身及分身理论的一个简短的总结。
-> 传统达尔文主义(Darwinism)
达尔文提出的原始进化论版本就是传统达尔文主义,它强调了基于生物可遗传性状在自然选择的作用下,经过微小缓慢的逐渐改变从而形成了更高级、复杂、适应性更好的物种。传统达尔文主义主要围绕两个主题展开:第一个主题是物种不断变化,缓慢地适应环境,且变化可以遗传;第二个主题是自然对物种做出选择,适者生存。达尔文由于时代的局限,在自己的书中对遗传机制的解释还相当晦涩模糊(他自己也承认对遗传如何起作用相当无知,在他那个年代因科技和知识的水平,无人不是如此)。
达尔文十分强调“微小”和“逐渐”的改变,把这视为他的进化思想中过渡方式的精髓,他在《物种起源》中说“倘使能证明有任何复杂器官不是经过无数的、连续的、轻微的变异而被形成的,那么我的学说就要完全破产。” 在1859年他给一位地质学家的信件中,他又一次强调说:“如果我的自然选择论必须借重这种(突然进化的)过程才能说得通,我将弃之如粪土。……如果在任何一个步骤中需要加上神奇的进步,那自然选择论就不值分文了。” 用达尔文的话来说大突变(突然进化)等于一个神迹,再极端一点来说大突变几乎与创造论没有什么分别。
-> 新达尔文主义 (Neo-Darwinism)
新达尔文主义产生于十九世纪末,创立者是德国生物学家、遗传学家奥古斯特·魏斯曼(August Weismann)。魏斯曼于格雷戈尔·孟德尔(Gregor J. Mendel)的遗传学说基础上提出了“种质”学说,该学说认为生物体可以分为“种质”和“体质”两部分,种质类似于我们现在说的基因或染色体,而体质相当于细胞质,也就是染色体以外的部分。魏斯曼把遗传学和自然选择学说结合起来,开创了进化论研究的新方向。美国遗传学家托马斯·摩尔根(Thomas H. Morgan)1917年提出“基因论”,把魏斯曼的种质发展为染色体上线性排列的遗传因子,即基因。对基因的研究揭示了遗传的变异机制,克服了达尔文进化学说对遗传机制描述的缺陷。在这个阶段对进化论的研究仍然停留在个体水平上,对自然选择在设想的进化中的可能重要作用没有统一的认识。
新达尔文主义强调基因突变是新物种形成的关键,用逐渐成熟的对遗传机制的理解再一次武装了进化论,是对传统达尔文主义的加强版变身。
-> 综合进化论(现代达尔文主义) (Modern Evolutionary Synthesis)
1942年英国生物学家朱利安·赫胥黎(Sir Julian S. Huxley)对遗传学、分类学和古生物学的研究成果进行了全面的综合,他的成果又由遗传学家费奥多西·杜布赞斯基(Theodosius G. Dobzhansky),动物学家恩斯特·迈尔(Ernst W. Mayr),和古生物学家乔治·辛普森(George G. Simpson)加以完善和发展,开创了综合进化论时代。综合进化论认为种群是生物进化的基本单位;认为突变和基因重组产生进化的原材料;自然选择可以保留有利变异和消除有害变异,从而使基因频率定向改变;隔离是物种形成的必要条件,如果缺少隔离,所有相似的物种混在一起随机杂交,就很难形成相对固定的形态。被隔离的群体有机会发展各自的性状,一旦这种性状稳定下来,后来就算隔离机制消失,那些新的性状也会散布开来。综合进化论在多门学科发展的基础上,继续保留了达尔文思想的核心,即自然选择。
综合进化论(现代达尔文主义)是在新达尔文主义的根基上进一步的经多学科整合的变身,也是现今进化理论的主体形态。
-> 中性演化理论(中性学说)(Neutral Theory of Molecular Evolution)
中性演化理论,又称“分子进化的中性学说”,认为分子水平上的大多数突变是中性或近中性的,自然选择对它们不起作用,这些突变全靠一代又一代的随机遗传漂变(生物繁衍时遗传机制,下段有解释)而被保存或趋于消失,从而形成分子水平上的进化性变化或种内变异。这个学说于1968年被日本遗传学家木村资生(Motoo Kimura)提出。木村通过对不同生物蛋白质的功能和结构的研究,发现突变大多是“中性”的,它不影响核酸和蛋白质的功能,对生物个体的生存既无害处,也无好处,“中性突变”通过随机的“遗传漂变”在群体里靠随机交配固定下来,学说认为在分子水平进化上自然选择不起作用(即自然不选择)。
现代遗传学研究指出,遗传漂变(源于DNA特定片段级别的编码整体交换)引起的等位基因频率在群体内的变化,不是由突变(源于DNA个别位点级别的编码随机变化)和自然选择造成的,而是单纯源于生物繁衍时遗传机制,即在配子形成时的减数分裂中四分体联会染色体片段重组。在小群体内部,漂变占主导地位,这会导致群体的遗传变异减少(某些等位基因会被快速固定,而某些则快速消失),即降低群体的杂合性(而突变的作用将提高群体的杂合性,但由于突变的频率很小,所以对改变群体的基因频率作用缓慢)。当种群变大时,漂变致使各种等位基因的频率趋于稳定。中性突变可以描述自然选择无法参与作用的性状,如人类头发的不同形状与颜色在种群中的形成和固定,对生物体的生存和繁殖不会发生影响,自然选择的作用也就无从谈起。遗传漂变也可以很好地解释遗传学中的单倍群概念,单倍群理论认为人类源自同一对原始父母,并通过迁徙和繁衍形成现在地球上不同地域和种族的人类群体分布。
与强调突变与自然选择作用的传统进化论不同,中性学说强调遗传漂变是分子进化的基本动力。换句话说,突变大多在种群中因遗传漂变而随机地被固定或消失,而不是通过选择才被保留或淘汰的。然而恩斯特·迈尔对木村的理论进行了猛烈的抨击,迈尔指出考虑到基因并不是自然选择的靶子,因此,所谓的中性进化就是一个毫无意义的概念。
中性学说是进化论的一个分身,也是对综合进化论强调自然选择作用的一次在进化机制上的反动。
-> 间断平衡论 (Punctuated Equilibrium)
间断平衡理论是由美国古生物学家尼尔斯·埃尔德雷奇(Niles Eldredge)与史蒂芬·古尔德(Stephen J. Gould),基于恩斯特·迈尔(Ernst W. Mayr)的异域物种形成,以及其它一些遗传理论在1972年所提出。在间断平衡的模型里,进化过程是由一种在短时间内爆发式产生的进化与在长时间稳定状态下的一系列渐变进化之间交替进行的过程。基因突变学说和地理隔离导致新物种快速形成是间断平衡论的理论基础。该理论强调大多数新种是从亲种地理分布的边缘被隔离的某个地区中的小种群中起源的,因为小种群中某些突变容易被选择保留,并随之产生生殖隔离而导致新的物种产生。该理论认为新种只能通过线系分支产生,只能以跳跃的方式快速形成(大突变);新种一旦形成就处于保守或进化停滞状态,直到下一次物种形成事件发生之前,表型上都不会有明显变化;进化是跳跃与停滞相间,不存在匀速、平滑、渐变的进化。
该理论认为,新种很快可以形成,只要在一始祖分布地域的边缘有一小组的个体与主流之间产生了分隔,就可以完成。自然选择的压力可能在某一生物种分布边缘地区向侥幸生存的群体发出最大的效力,并且幸免死亡而存留的异种很快就可以在这数目很小被分隔的群体中散布。这样,一个全新的生物种就可能在大群体的边缘产生,并且不留下任何化石的痕迹,因为化石主要是从大群体中的动物遗留下来的。这样新种就好像在化石记录中突然出现,然后渐渐分布到整个始祖种群占领的地区。
间断平衡理论有两个重要支撑点:首先,化石表明物种呈现明显的稳定性,在地质记录中出现和消失时的外形几乎相同,没有出现达尔文所说的连续变化;其次,新物种的出现是突然事件,而且一旦出现,就已相当完备,没有显示出进一步的修改(参看第三节中古尔德对化石形态的稳定性和突然出现的现象的总结)。这些与达尔文的渐变进化模型完全抵触。
恩斯特·迈尔认为所谓的大突变过程根本就不会存在,因为一个个体的基因是一个协调、平衡的系统,这个系统是在几百万年的时间里,通过一代代的自然选择,最终形成并协调好的;既然已经知道绝大多数基因位点上的潜在突变会产生有害或致死的效应,一次重大的突变所导致的整个基因型的大震动又怎么能产生出能够繁衍下去的个体呢?到哪儿去发现这样从一个大突变过程产生出的未成功成活的数量巨大的变种呢?
间断平衡理论是进化论的另一个分身,是进化论学者对化石中间形态缺失的困惑的唯一一次富有勇气的尝试。间断平衡理论(其实质是进化论和创造论之间的中间骑墙路线)无疑是对传统达尔文渐变学说的最大的反动。渐变论的忠实支持者,当代综合进化论的旗手人物、原牛津大学的动物行为学家理查德·道金斯(Richard Dawkins)评价大突变机制说“你可以把《圣经》说人是用泥土造的看作是大突变”,以表达对大突变机制的厌恶。间断平衡论所要求的在小种群内部的大突变同样面临迈尔指出的“重大的突变所导致的整个基因型的大震动”发生和存续的可能性和机制的问题。而且在现实观察中所见生物的大突变都是畸形,从未见有对生物的生存繁衍有帮助的大突变出现,更勿论整套的器官和身体系统的出现了。
-> 所谓的“自私的基因”
谈到大名鼎鼎的理查德·道金斯,这里有必要介绍一下他的所谓的“自私的基因”,这个概念是他在1976年他的第一本书《自私的基因》中提出的,即进化的过程是被基因驱动的,基因的目的只有一个,那就是不断地,更多地复制自己;作为进化的单元,基因自私并且只对自己的生存和繁殖感兴趣。为此古尔德辛辣地指出,道金斯的基因选择机理是把鸡看成是一只鸡蛋制造另一只鸡蛋的通道,那只鸡只是一个无足轻重的工具。古尔德认为他找到了所谓“自私的基因”的致命的缺陷:自然选择无法对基因直接施加影响,即自然无法“看见”基因,所有的基因都戴着层层的面纱深藏在身体之中,而自然接触的只是身体。古尔德得出结论:单独来看,每个基因的自私性与否是没有意义的,自然选择只能发生在个体水平上,而不是基因水平上。
道金斯的这部书不是学术著作,而是一部披着科普外衣的伪科普作品。作者运用自己天马行空的想象力,赋予了基因以某种“意识”,然而他所谓的“自私的基因”似有为某些人类个体极端的利己主义背书之嫌。所谓的“自私的基因”可能会带给某一类受众以特有的、诗意般的美感,但它从任何意义上说都不是一种理论。道金斯的其它著述也能体现出他的逻辑思维混乱,其人缺乏合格的人文及哲学修养,而以卖弄噱头为看点,在他的书中他羸弱的思辨能力和他所试图论述的许多宏大议题时常显得不相匹配。进化论发展到今天,道金斯这种学品的人得以成为旗手人物,是进化论的悲哀。
通过以上简短的介绍,读者应该可以看出在进化论阵营的内部,对种间宏进化机制如何起作用众说纷纭,莫衷一是。在尝试找寻真理的崎岖的雾霾重重的山路上,他们跌跌撞撞、步履蹒跚,艰难地尝试自圆其说,然而一次次的努力都显示他们离朝思暮想的成功还相去甚远。
这里提供一个表格,对从传统进化论以来的各个变身、分身理论的在解释物种间宏进化机制时的困难做一小结:
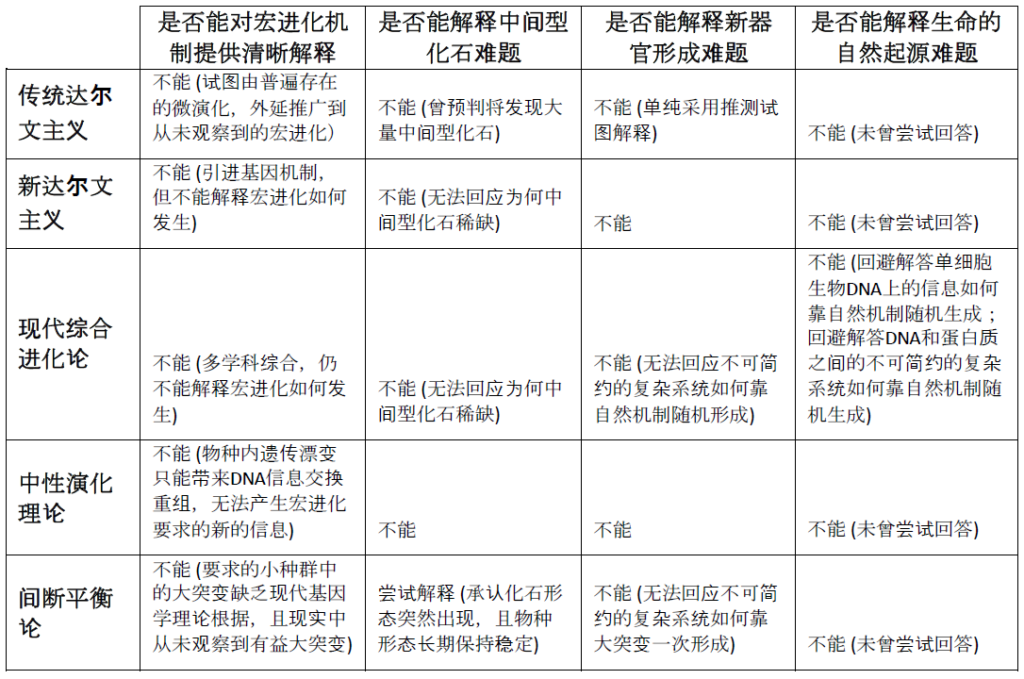
有一点需要特别指出的是,由于自十九世纪后半叶开始,进化论逐渐在科学界成为生物学压倒性质的主流理论,这就不奇怪为什么我们看到后来很多科学家如摩尔根、杜布赞斯基、古尔德等都以进化论作为信仰框架(在第十节详细讨论)在相关的领域进行科学研究。他们的很多研究成果如基因的遗传机理等都是著有成效的,对人类的科学进步做出了贡献,然而这些学科研究的成果还远不足以解释宏进化的发生机理,因而这些产生成果的事实并不能作为进化论的核心主张已经成功的证据;相反,基因学的研究显示生物的基因在物种的层次上具有强大的稳定性(Genetic Homeostasis),这与生物在化石中保留的远古形态和现今形态的极度相似的观察是一致的。其实公允地说,即使是达尔文本人,从他写作的风格也可以看出他是一个试图做到尽量学术严谨的著作者,从同时代他周围的人给他的评价也显示他是一个公认的绅士;他的理论极有可能是错误的,这导致他也许不是像牛顿那样伟大的科学家,但是不能因此否定他作为人的其它价值。还可以更进一个层次说,其实连错误的理论都有它的价值,两相对比,正确的理论会愈加显为正确,《圣经》所宣扬的神不是也说“你们来,我们彼此辩论” (”Come now, and let us reason together”) 吗?
第十节. 对进化论逻辑的再思考
在本文的最后一节,让我们来分析一下进化论所使用的逻辑,以期使读者进一步理解进化论推理时可能存在的谬误。
首先我们要看到,达尔文在他理论形成的根基上,就相当有可能已经犯了以偏概全、无限延申的逻辑错误。他认为在物种内部随处可见的物种性状的无方向性变化,可以用来解释他期望的物种间由少到多、由低到高的所谓的进化(第二节有详细讨论)。而根据经验,我们已经确知,在这个世界上,微小的量变不一定导致飞跃性的质变;根据已知的观察和证据,种间的宏进化应该从未发生过。这就难怪由他肇始的遗传变异加自然选择的进化论在解释中间过渡类型化石缺乏、新器官形成机制、和生命从无到有的机制等方面举步维艰。
进而我们要看到,在进化论的语境下,进化论者有严重的循环论证之嫌。读者如果在维基百科里查词条“比较解剖学”,就会读到:“比较解剖学是在解剖学角度中比较生物的相似与差异处的一门研究,其通过比较的方法对比不同机体的结构特征,并观察分析其相互间的异同,从而了解生物进化的发展规律。” 对比较解剖学里的重要概念“同源结构” 其词条解释为“不同物种中的相似结构(身体部位、器官等),因为物种有共同起源。它们可能用以执行相同的功能。” 好了,停到这里仔细看:进化论以“相似”做为推理的前提,又用“相似”做为推理的结论,不是吗?—— 最初进化论是以生物的相似性,推出生物有共同祖先,生物是进化来的;在解释生物的相似性时,进化论者又说因为生物是进化来的,所以生物有相似性。进化论者不过就是在说“因为A现象所以B成立”,然后在同样的语境下又说“因为B成立所以有A现象”,这前后两句话互相指引互相支持,彼此作为彼此为真的论证,然而这在逻辑上是不具备实质论证意义的,它只是在自己铺陈好的语境里作循环自证,这是一种诡辩术。
进化论提出“因为人和猿有共同祖先,所以人和猿有很多相似的特征。”;进化论又陈述“因为人和猿有很多相似的特征,所以人和猿有共同祖先。”;这一前一后两句话,彼此互为逆命题。然而逻辑上一个说法和它自己的逆命题两者是否都同时为真,是没有彼此的直接联系的,这就好比说“因为今天是爱丽丝的婚礼,所以她穿了婚纱。”,那么它的逆命题“因为爱丽丝穿了婚纱,所以今天是她的婚礼。”就一定为真吗?
对进化论的逻辑的分析还可以更进一步,而且是这节分析的更为关键的一步。如前对进化论的发展的介绍,进化论在最初由达尔文发表的时候,是使用不同物种在形态和结构上的相似为主要依据而导出不同物种是从共同祖先进化来的;但进化论面对中间型化石缺乏的困境,在最近半个多世纪依赖分子生物学和基因遗传学的发展,它又转而诉诸相似生物在DNA编码和蛋白质结构方面的相似性来支持其生物进化观点,很多读者对这些都已经不陌生了。但是不知读者们意识到没有,进化论的死敌——创造论(这里指“特创论”,即认为从未出现过生物种间的进化,生物种类是创造者按“各从其类”的原则一次就创造完成的),对相近生物在宏观层面的形态和结构上的相似性,以及它们在微观层面的DNA编码和蛋白质结构方面的相似性,比如说“人和猿有很多相似的特征”这一点,也可以有一摸一样的预期。
如果这个星球上的生物存在一位创造者,作为读者的你,会设想那位创造者用什么样的方式来创造所谓的“自然界”中林林总总的生物呢?创造者会像一个艺术家那样把其所有的作品都创造得独一无二,还是会像工程师那样用尽量少的有效设计的范型而推广应用到所有需要相似功能的生物身上呢?我们认为创造者可以选择把所有的生物都创造得与其它生物不同,也可以选择把很多生物都创造得相似,这是创造者所天然具有的自由。
相似的器官和身体结构一定是要靠进化才能得来的吗?可能存在的创造者为什么不能选择就把它们创造得相似呢?黑猩猩的手臂和人的手臂无论从结构还是功能都高度相似,如果存在一位创造者,我们认为其可以选择把它们创造得相似,以期用最高的设计效率实现应用效果的最大化,进而实现自己的创造目的,这是工程(engieering)中最基本的思想。如果我们在亨利·福特的汽车的实验型和后来量产的商业型之间发现很多相似,相信没有人会感到惊异,因为从实验型到商业型都是出自同一个工程师的设计(即“同源设计”)。当然,从生物界精彩纷呈的角度来看,如果读者仔细审视“所谓的”自然界(如果存在一位创造者,那么“自然”就不过是“超自然”的创造)中那一件件带给人美感的被造物,那么那位可能存在的创造者不但是一位最高超的工程师,祂同时也是一位最卓越的艺术家。
在生物DNA编码和蛋白质结构方面的相似性也同样可以用可能存在的创造者解释得通。有计算机编程知识的人都知道,现代编程的极其重要的理念之一就是强调代码的模块化封装和重用,以期实现潜在的出错率的最小化和编程效率的最大化。如果一段程序代码可以简单、安全、高效的实现一种特定的功能,那么这段代码就会被编程者重复使用。如果在黑猩猩的基因中发现其指导血红蛋白生成的编码和人类基因中指导血红蛋白生成的编码相似(或完全相同),就能证明黑猩猩和人类有共同祖先吗?如果存在一位创造者,其显然有自由决定在两个不同的生物之间使用相似的(或完全相同的)DNA编码来指导血红蛋白的表达。
由此可以看出,由一位可能存在的创造者,同样可以解释相似生物在宏观形态结构方面的相似,和在微观DNA编码及蛋白质结构方面的相似。进化论以自己之一厢情愿,使用这两点作为其假说成立的依据,其逻辑根基总显得不那么坚实。进化论者可能还没有意识到,他们孜孜不倦的在生物结构和DNA编码相似性方面的研究诠释,可以完美地被他们的敌手创造论者原封不动地搬来用以佐证那位可能存在的创造者的存在。进化论者诉诸那个观察不到(从未被观察到,将来也极有可能不会被观察到)的物种间进化机制来解释这个星球上所有物种的存在,而创造论者用以解释同样现象时诉诸的是那位难以观察到的超自然的创造者,进化论者显然是凭借无比巨大的、主观的信心(至少我们看不到他们是凭借有说服力的证据)才去质疑创造论者的吧。
讲到DNA上的编码,我们已经知道DNA上的编码本质上是信息,其上的含有A、T、G、和C碱基的核苷酸是信息的载体(物质),而A、T、G、和C编码的总体在信息载体的层次之上,进而保留和传递了指导生命生存和延续的信息(非物质)。正如印在纸上的这句话中”All aspects of the evidences for evolution theory have been weighed on the scales and found wanting.”,印刷字符的油墨和承载油墨的纸张是信息的载体(物质),而在油墨和纸张的层次之上,由字母空格等字符在总体上传递了一个信息(非物质)。
现在笔者想请读者思考这样一个问题:载体(核苷酸、油墨和纸等)是物质的、出于自然的,但非物质的信息是从哪里来的?你期望上段中那句由字母和空格组成长为100个字符的信息靠自然界随机经历亿万年生成吗?恐怕自然界经历一亿个亿万年也无法形成一个这样的信息,这样的信息(哪怕是这样短短的信息)在宇宙中唯一可能的出处是“智能”。下图给出一个对比,左侧是自然的“创作”——呈现随机低序的物质组合,右侧是智能的创作——在物质的层次之上传递具有特殊复杂性(Specified Complexity)的信息。

这个星球上迄今最为庞大复杂的代码系统之一应该就是微软Windows操作系统的源代码;智识正常的人应该都不会否定,自然不但无法靠随机生成这个级别的有意义的源代码,而且连轻微地改动源代码的能力都不具备。设想将载有Windows操作系统源代码的计算机放在一只猴子的面前,让猴子在键盘上随机(模仿自然)敲击若干下,然后将程序重新编译,有任何人会期望这样经过随机修改的程序还能执行吗?而进化论一直给所有人灌输的概念就是,自然有能力不断靠随机修改DNA上的信息而使系统增加功能(先不讨论进化论不愿多谈的信息最初是怎么产生的问题),这和让我们期望经猴子随机改变后的代码又为Windows添加了新的功能有任何本质的区别吗?随机的机制不可能产生真正具有意义和功能的信息,这和近半个世纪以来科学对各种生物(从最低级的细菌到最高级的人)的DNA上信息的研究观察的结果也完全一致,那就是我们只观察到了DNA上由于突变可以发生信息的丢失或轻微的改动,以及由于生殖细胞在减数分裂(对有性生殖的生物来说)时的联会而发生的有限范围内的信息的交换重组,而没有任何研究者观察到DNA上产生出新的具有意义和功能的信息且能被生物传承下来。换言之,地球上的所有生物体内的DNA上的信息在物种的范围内都具有极其让科学惊叹的稳定性(Genetic Homeostasis)。
微软公司的创始人比尔·盖茨在评论DNA编码的复杂程度时说:“DNA就像是计算机程序,但比任何已经创造出来的软件都要先进得太多太多。” 对应哪怕是单细胞生物体内的DNA上的信息,Windows程序中的源代码级别的信息也只能算作婴儿科;如果有进化论者可以说服笔者哪怕是Windows操作系统级别的信息系统可以只靠自然界随机形成,那么笔者愿意立时抛却对进化论的所有疑惑,和所有的进化论者一样,拜倒在这个作为“现代科学基石”的、带给人类空前“思想自由”的、“伟大”的“科学”理论脚下。
2001年世界范围内许多教研机构(包括欧美众多知名大学和科研实体)的多学科的众多的科学家联名发布了一个名为《对达尔文主义的科学异议》(《A Scientific Dissent From Darwinism》)的声明,迄今为止在这个声明上签字的科学家已经超过了一千名。该声明明确表示:“我们对随机变异和自然选择能够解释生命复杂性的观点表示怀疑。应该鼓励对达尔文学说的证据进行谨慎的检验。”
讨论至此,作为此篇文章的结论,如果我们说进化论不像科学,而更像一个信仰系统,相信读者们已经不会太感到意外了。科学(自然科学)的基本特征是可以解释观察到的现象,并能对未来可能发生的现象做出预判。爱因斯坦提出的相对论,可以解释高速运动的系统中时间的膨胀,也能对光线经过大质量天体时产生弯曲做出准确的预判,这就是为什么几乎没有人反对在科学发展的现阶段,相对论是科学的,或至少是接近真理的。进化论试图解释生物在遗传和自然选择的作用下由低等到高等的所谓的进化的论点,其论据的采用和论证的过程都相当不具有说服力(即极度缺乏解释力),它预判的所谓的“大量的中间环节化石的发现”,从达尔文到现在来看也是遥遥无期。由达尔文肇始的进化论迄今为止的解释力和预判力可谓积贫积弱,由此看它不是严格意义上的科学,相信它的人是需要相当程度的“信心”(faith)的。(信心:基于领悟感受而不是全然可检验的证据的,强烈坚定地信入某事物。)
进化论者无不是无意或有意地站在自然主义哲学的预设立场来秉持、诠释和推广他们心中的进化论,而不是严格地凭借科学所要求的坚实的证据和缜密的推理,在此仅举出两例。
第一例. 达尔文在1859年写给一位地质学家的信(与上节所述为同一封信)中谈到自然选择说:“但是我对它有坚定的信心,因为我无法相信如果它是错的,它竟可以解释(至少在我看来它能够解释)那么多层次的事实。” 在这里读者能不能体会到达尔文“相信”他的进化论是正确的,多少是要凭借他“坚定的信心”(firm faith)呢?
第二例. 古尔德在他的文章《进化作为事实和理论》中说: “无论科学家怎样根据不同学说争辩应当如何对事实作正确的解释时,事实仍不改变。爱因斯坦的引力理论代替了牛顿的理论,争辩期间苹果并未因此悬在半空期待结论。同样,人类从似猿的祖先进化而来也是事实,无论是照达尔文提出的机制或其它什么机制,这仍然有待决定。” 然而我们要抗议说,连一岁的孩童都能观察到苹果落地,但从笔者在本文中所有的分析和列举的证据来看,我们很难被说服而认为人类与猿类有同一祖先,古尔德是凭借什么样的“信心”得出“人类从似猿的祖先进化而来也是事实”的结论来的呢?从古尔德的这段话中,读者是不是也像笔者一样读出了他是凭着对进化论的“信心”,在进化机制远未能确定的前提下,断言“进化是事实”的呢?
这样只凭借自己的哲学世界观立场而忽略科学的客观证据的现象在进化论者中间比比皆是。其实不难看到,在自然主义的立场下,自然选择和进化是立场持有者的唯一的选择,正如达尔文同代的对其理论最坚定的支持者托马斯·赫胥黎(Thomas H. Huxley)面对他人的对进化论缺乏证据支持而提出的疑问时,就会毫不犹豫地反问道:“你还有什么其它的选择呢?”
如果我们在这整篇文章中的分析基本正确的话,由于进化论无论是在解释地球上生命的出现,还是在所有物种的形成方面,都几乎不得不借助奇迹,这和创造论宣扬的神迹在本质上并不是科学和信仰的区分,而是两个不同的信仰之间的区分。进化论是关乎“自然的创造”的信仰,而创造论是关乎“超自然的创造”的信仰。
从需要“信心”这个意义上来说,进化论和创造论相似,都是信仰系统,其持有者须凭借自己的自由意志(free will),“选择”对其相信,而无法在人的层次上对这种选择提供终极的、确定的理性支持。从信仰系统的方面来说,进化论(究其本质)和创造论一样,都需要持有者自愿地靠信心(faith)才能最终抵达其核心;换言之,单单凭借人的理性(rationality)无法最终抵达。但是理性显然也有正确的(合理的)和不正确的(不合理的)之分;进化论不是科学,它是一个用科学的外衣包裹的、与自然主义哲学相互倚重的信仰;在科学的范畴下,进化论充其量是一个假说,而且是一个高度疑似不具科学性的、使用错误理性的假说,是一个必须靠相信它的人联手发布声明来维持的假说;由于进化论严重缺乏坚实的证据支持,可能它将永远也只能是个假说。进化论,百年迷雾,千疮百孔,岌岌可危,笃信它的人应该重新检视一下它的合理性了。
在文章的最后,笔者要借用一位曾经行走在这个星球上的“人”的话作为结束送给有耐心读到这里的读者(同时也再一次提醒笔者自己):“你们将明白真理,而真理将使你们自由。” (”You will know the truth, and the truth will set you free.”)
_______________________________________________________________
附录1. 本文章主要参考书目及资料来源网址(排名按中文拼音及英文字母顺序)
- 百度百科 https://baike.baidu.com/
- 《长颈鹿:又一例不可简化的复杂性》 https://www.chuangzaolun.com/
- 《达尔文的黑匣子:生化理论对进化论的挑战》 迈克尔・J・贝希 (邢锡范译), 中央编译出版社 1998年
- 《飞行:不可简化的复杂性》 https://www.chuangzaolun.com/
- 《黑猩猩和人类基因组的DNA序列相似性达到99%》 https://www.cas.cn/xw/kjsm/gjdt/200906/t20090608_632638.shtml
- 《进化论的死穴》 https://www.youtube.com/watch?v=v38JhzIgBjE
- 《盲眼钟表匠》 理查德・道金斯 (王道还译), 中信出版社 2014年
- 《其实你不懂进化论》 史钧,世界图书出版社 2020年
- 《审判达尔文》 詹腓力 (钱锟译), 中央编译出版社 2006年
- 维基百科 https://www.wikipedia.org/
- 《物种起源》 查尔斯・达尔文 (周建人等译), 商务印书馆 1997年
- 《依据DNA编码的物种起源学 – 寻找科学亚当与科学夏娃》 张维克 宋国平, Life and DNA Press 2021
- 《A Scientific Dissent From Darwinism》 https://dissentfromdarwin.org/
- 《Chimpanzee and Human Y Chromosomes Are Remarkably Divergent in Structure and Gene Content》 《Nature》 Jan.2010
- 《Citrate Death Spiral》 Michael Behe,https://evolutionnews.org/2020/06/citrate-death-spiral/2020
- 《Construction and Analysis of a Human-Chimpanzee Comparative Clone Map》 《Science》Jan.2002
- 《Darwin On Trial》 Phillip E. Johnson, InterVarsity Press 2010
- 《DNA replication (DNA复制机制)》 https://www.youtube.com/watch?v=VisJOA-7Ef0
- 《Expelled: No Intelligence Allowed (full movie)》 https://www.youtube.com/watch?v=V5EPymcWp-g
- 《From DNA to protein – 3D》 https://www.youtube.com/watch?v=gG7uCskUOrA
- 《Genomic monkey business—estimates of nearly identical human–chimp DNA similarity re-evaluated using omitted data》 Jeffrey Tomkins and Jerry Bergman,https://creation.com/ Oct.2012
- 《How Radiometric Dating Works: Relative not Absolute Ages – Dr. Andrew Snelling (Conf Lecture)》 https://www.youtube.com/watch?app=desktop&v=z1lBdLVyzzo&t=26m52s 2018
- 《Initial sequence of the chimpanzee genome and comparison with the human genome》 《Nature》 Sep.2005
- 《On The Origin of Species by Means of Natural Selection, Or The Preservation of Favoured Races In The Struggle For Life》 Charles Darwin, New York: D. Appleton And Company 1860
- 《Probability of a Single Protein Forming by Chance》 https://www.youtube.com/watch?v=W1_KEVaCyaA
附录2. 本文章所用示意图表来源网址